Том 2
Эволюция природной среды
Морские бассейны
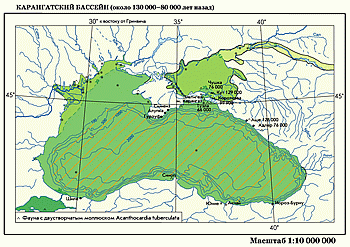
Карангатский морской бассейн занимал Черноморскую котловину во время максимума последнего (микулинского) межледниковья.  Уровень моря был на 6–8 м выше современного. Береговая линия продвинулась вглубь континента по долинам Днепра и Южного Буга.
Уровень моря был на 6–8 м выше современного. Береговая линия продвинулась вглубь континента по долинам Днепра и Южного Буга.  По Дону и Манычу морские воды проникли вверх до устья Калауса. Они заливали дельту Дуная и залив Сиваш. Карангатский бассейн имел более широкую связь со Средиземным морем (Тирренский бассейн). Его соленость была выше современной и достигала 30‰. В долинах рек и лиманах морские воды опреснялись до 15–20‰. В закрытых лиманах сохранялись солоновато-водные условия для развития каспийской фауны — Didacna, Monodacna, Dreissena. Глубже 100–200 м до самого дна господствовали безжизненные условия сероводородной зоны. На дне шельфа и в заливах Карангатского моря обитала донная фауна моллюсков средиземноморского типа
По Дону и Манычу морские воды проникли вверх до устья Калауса. Они заливали дельту Дуная и залив Сиваш. Карангатский бассейн имел более широкую связь со Средиземным морем (Тирренский бассейн). Его соленость была выше современной и достигала 30‰. В долинах рек и лиманах морские воды опреснялись до 15–20‰. В закрытых лиманах сохранялись солоновато-водные условия для развития каспийской фауны — Didacna, Monodacna, Dreissena. Глубже 100–200 м до самого дна господствовали безжизненные условия сероводородной зоны. На дне шельфа и в заливах Карангатского моря обитала донная фауна моллюсков средиземноморского типа  (Acanthocardia tuberculata, Paphia senescens, Mactra corallina, Apporhais pespelicani и др.), но более обедненная. В толще воды обитали планктонные фораминиферы и нанопланктон (коколитоф o риды). Температура воды была выше современной, о чем свидетельствуют устрично-серпулево-водорослевые биогермы с теплолюбивыми видами моллюсков, отсутствующих сейчас в фауне Черного моря, но обитающих в Средиземном море.
(Acanthocardia tuberculata, Paphia senescens, Mactra corallina, Apporhais pespelicani и др.), но более обедненная. В толще воды обитали планктонные фораминиферы и нанопланктон (коколитоф o риды). Температура воды была выше современной, о чем свидетельствуют устрично-серпулево-водорослевые биогермы с теплолюбивыми видами моллюсков, отсутствующих сейчас в фауне Черного моря, но обитающих в Средиземном море.
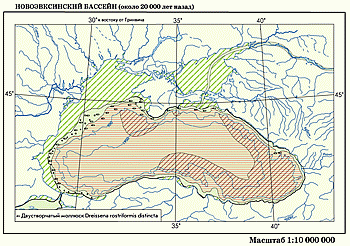
Новоэвксинский бассейн. Во время максимума последнего оледенения образовался полуизолированный регрессивный проточный Новоэвксинский бассейн полупресноводного типа. Его уровень во время пика регрессии опускался на 100–110 м, то есть ниже порога в Босфоре. Площадь бассейна была на 100 тыс. км2 меньше современной и составляла всего 360 тыс. км2 .
Вслед за понижением уровня моря осушались большие участки шельфа, особенно в северо-западной части, где обширные пространства были заняты сушей, пересеченной долинами Дуная, Днепра, Днестра. Азовского моря из-за его мелководности не существовало, его пересекали продолжения долины Дона и Маныч-Керченского пролива, по которому сбрасывались воды Хвалынского бассейна из Каспийской котловины.

В связи с прекращением притока вод из Средиземного моря Новоэвксинский бассейн трансформировался в полупресноводное озеро-море с минимальной соленостью (до 6–7‰). Воды из него сбрасывались в Мраморное море по проливу Босфор, в донных отложениях которого выявлены каспийские моллюски Dreissena rostriformis .
При таких условиях в составе фауны сохранялись только наиболее опресненные виды каспийского типа — Dreissena rostriformis, Dr . polymorpha, Monodacna с aspia, Adacna plicata и другие, но без Didacna . Для этого моллюска здесь была слишком низкая соленость. Новоэвксинский бассейн представлял собой звено каскада евразийских бассейнов от Аральского моря до Средиземного.
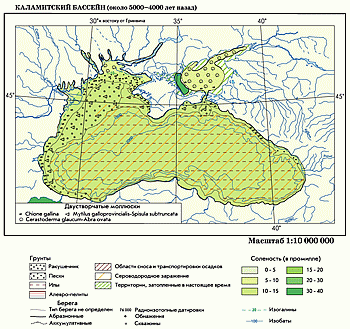
Каламитский полуморской бассейн. На рубеже раннего и среднего голоцена Черноморская котловина заполнилась солеными средиземноморскими водами. Уровень океанической трансгрессии превысил порог стока в Босфоре и соленые воды проникли в Новоэвксинский бассейн. Морские воды затопили почти все пространство современного шельфа, вновь восстановилось Азовское море. Море не только достигло современной береговой линии, но проникло по долинам рек на десятки и сотни километров вверх по течению. Этому способствовало позднеледниковое переуглубление долин крупных рек, которые ныне в значительной степени заполнены голоценовыми осадками.

Резко возросла соленость до 15–20‰ и ее тип изменился с континентального на океанический, быстро потяжелел изотопный состав вод, началась стратификация водных масс по солености и формирование зоны сероводородного заражения.

Особенно большие изменения претерпела биота. Пресноводные организмы на большей части акватории были уничтожены морской трансгрессией, а их остатки, оттеснены в пресноводные лиманы и эстуарии. Основная акватория шельфа была быстро занята морскими видами и биоценозами, проникшими из Средиземного моря. На внутреннем шельфе преобладали биоценозы Mytilus galloprovincialis — Spisula subtruncata на алевритово-ракушечных грунтах. Внешний шельф и прибрежная зона с песчано-алевритовыми грунтами были заняты биоценозом Cerastoderma glaucum — Abra ovata . Наконец, по периферии полуморского бассейна в устье Днепра, а также в мелководном Азовском море сформировались опресненные биоценозы Chione gallina .
В глубоководной впадине ниже 150–200 м находились анаэробные водные массы, зараженные сероводородом. Безжизненное дно впадины устилали илистые отложения. На восточном континентальном склоне усилились склоновые процессы, связанные с оползанием осадков, мутьевыми потоками и образованием турбидитов — отложений турбулентных мутьевых потоков.
Каспийское море
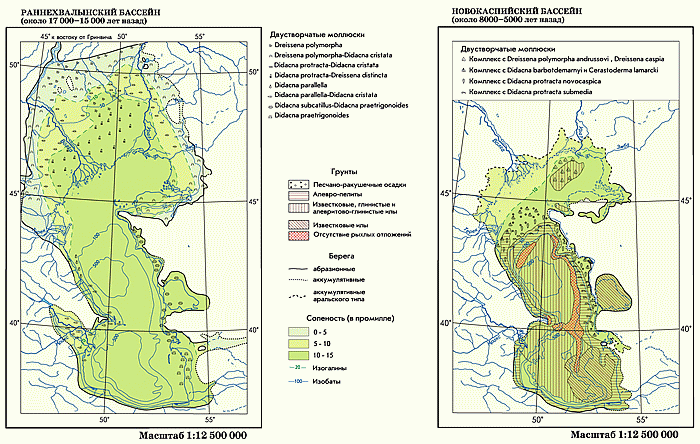


Раннехвалынский бассейн — самый большой по площади бассейн в плейстоценовой истории Каспия — сформировался на стадии дегляциации последнего оледенения Европы. Площадь его акватории достигала 800–900 тыс. км2, что в 2,5 раза больше современного Каспия. В результате очень быстрой трансгрессии уровень Раннехвалынского моря поднялся почти на 200 м и достиг абсолютной отметки 48 (50) м. На севере береговая линия продвинулась до подножий окружающих море возвышенностей (Ергени, Приволжская, Общий Сырт, Устюрт и Ставропольская). По долинам рек образовались длинные заливы типа лиманов, а по долине Волги — самый большой эстуарий длиной до 800 км, воды которого достигали устья Камы и современного города Чебоксары. На востоке воды этого бассейна покрывали закаспийские низменности и частично Каракумы и сливались с Арало-Сарыкамышским озером-морем в единый бассейн.
Соленость моря из-за его проточности была несколько ниже современной — около 10–12‰. Соленые воды Раннехвалынского бассейна сбрасывались через Зунда-Толгинский пережим в Манычскую впадину и далее по дну Азовской впадины в Новоэвксинский бассейн. Расход сбрасываемых вод достигал 10–50 тыс. м3/ c ек. Раннехвалынское море населяли эндемичные каспийские виды моллюсков солоновато-водного типа — Didacna trigonoides, D . protracta, D . ebersini, D . parallela, Monodacna caspia, Dreissena rostriformis и др.


Новокаспийский бассейн. В послеледниковое время размеры, уровень и соленость Каспийского бассейна приблизились к современным параметрам. Колебания уровня стабилизировались на абсолютных отметках от –20 до –30 м. Береговая линия была также близка к современной, бассейн оставался бессточным солоновато-водного типа.
Во время дербентской регрессии ( VIII – IX вв.), предшествовавшей последнему трансгрессивному пику, уровень моря снижался до отметок –32...–34 м.

Донные моллюсковые биоценозы Новокаспийского бассейна по составу были близки к современным. Наиболее опресненная акватория Северного Каспия вблизи дельт Волги и Урала была заселена комплексом пресноводных моллюсков с преобладанием Dreissena polymorpha andrussovi — Dr . с aspia. Основная шельфовая акватория Среднего и Южного Каспия была заселена биоценозом Didacna barbotdemarnyi — Cerastoderma lamarcki ( Cardium edule). Более глубоководные участки Среднего и Южного Каспия (глубже 30–50 м) были заняты биоценозом Didacna protracta novocaspia, а самые глубоководные — биоценозом Didacna protracta submedia .
Соленость вод, примыкающих к дельтам Волги, Урала и Терека, была менее 10‰, а соленость основной акватории превышала 10‰.
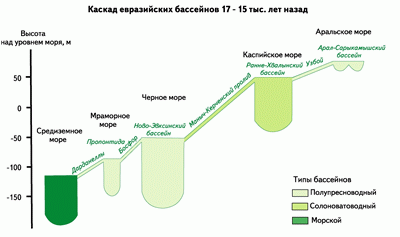
В результате таяния материковых льдов и вечной мерзлоты последнего (Валдайского) оледенения во внутренней Евразии образовалась система связанных между собой водоемов, объединенных в каскад евразийских бассейнов от Арала до Средиземного моря. Каскад начинался с Арало-Сарыкамышского бассейна, заполнявшего две неглубокие (до 100 м) котловины — Аральскую и Сарыкамышскую впадины площадью акватории около 100 000 км2 и объемом до 1500 км3. Соленость вод в них не превышала 5‰. Через пролив Узбой длиной 600 км эти воды стекали в огромное Раннехвалынское озеро-море, глубина которого ( 1120 м) превышала глубину современного Каспийского моря, а объем вод (135 000 км3) был больше современного в два раза. По Маныч-Керченскому проливу длиной около 1000 км воды Хвалынского моря сливались в Новоэвксинское озеро-море, а далее через пролив Босфор (длиной 45 км) осуществлялся односторонний сток в древнее опресненное Мраморное море (Пропонтиду), по солености вод и составу флоры и фауны близкое к Новоэвксинскому бассейну. Наконец, через пролив Дарданеллы воды этого каскада сливались в Эгейское море. По своим масштабам озерно-морская система каскада евразийских бассейнов не имеет сейчас аналогов на Земле: площадь ее акватории достигала 1,5 млн км2, объем воды составлял 700 тыс. км3, расход сбрасываемых вод превышал 60 тыс. м3/сек.
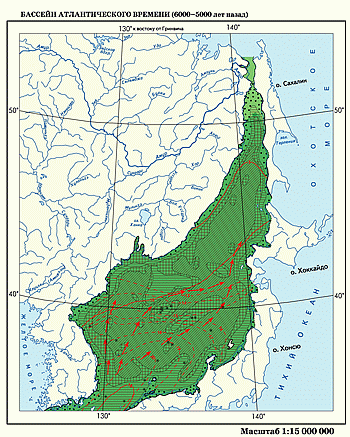
Бассейн атлантического времени. Очертания береговой линии в эпоху климатического оптимума голоцена незначительно отличались от современных, однако связь с окружающими морями была несколько большей.
Режим осадконакопления определялся сочетанием трех важных для седиментотенеза факторов: общим увеличением температур, оптимальным водообменом с окружающими морями и Тихим океаном, а также резким уменьшением поступления терригенного обломочного материала на шельф и в глубоководные части Японской впадины. Первые два фактора действовали сопряженно, так как одновременно с планетарным потеплением климата температура поверхностной водной массы в Японском море возрастала за счет притока теплых вод. Поэтому в южной и центральных частях бассейна накапливались алевропелитовые или пелитовые илы, обогащенные раковинами планктонных фораминифер. Усиленный водообмен с Восточно-Китайским морем хорошо диагностируется по составу глинистых минералов. Преобладание среди последних гидрослюд (особенно в зоне действия Цусимского течения) явно отвечает активному поступлению терригенного материала, выносимого великими реками Восточной Азии в омывающие ее моря и перераспределяемого затем течениями. С более активной циркуляцией водных масс связано и общее уменьшение пелитов в составе донных отложений Японского моря.
Подъем уровня моря до отметок, близких к современным, привел к восстановлению связи Японского моря с акваториями Охотского, Восточно-Китайского морей и Тихого океана. Это привело к превращению Японского моря в нормальный морской бассейн. Активное поступление теплых морских вод на фоне общепланетарного потепления климата вызвало замену субарктической фауны фораминифер в центральной части Японского моря бореальной, а на юге бассейна —субтропической. В эпоху климатического оптимума среднегодовая температура поверхностных вод превышала современную на 1–2°С, а в прибрежных и мелководных частях на 3–5°С (по палинологическим данным) или на 5–8°С (по результатам изучения бентосных фораминифер). На сильное потепление климата и повышение температуры водных масс Японского моря указывает смещение ареалов субтропических и тропических видов диатомей почти на 1000 км к северу от современной границы их распространения.
Значительное распространение в прибрежной части севера Японского моря широколиственных и кедрово-широколиственных лесов, в настоящее время сопряженных с малоледовитыми в зимнее время акваториями, служит косвенным признаком смещения южной границы плавучих льдов в северную часть акватории Татарского пролива.
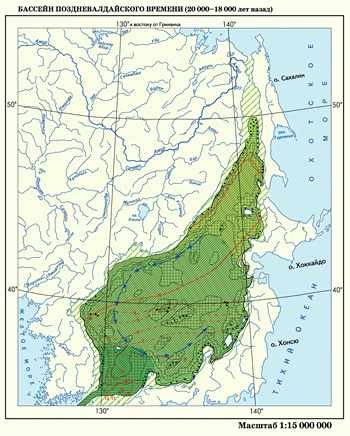
Бассейн поздневалдайского времени. Понижение уровня Мирового океана в эпоху максимального похолодания на 130–140 м привело к резкому изменению береговой линии и к относительной изоляции Японского моря. Почти все проливы, кроме Цусимского, были осушены. На месте Татарского пролива остался глубокий залив, исчезли мелководные проливы Невельского, Лаперуза, Сангарский и Восточно-Цусимский. Японское море трансформировалось в полуизолированный бассейн с единственной связью через Западно-Цусимский пролив.
В связи с поступлением с севера амурской пресной воды образовался значительный градиент солености: от 20‰ в Татарском заливе до более 30‰ в Цусимском проливе. Аналогичным образом проявился градиент среднегодовой температуры воды: от 5°С в Татарском проливе до 12°С в Цусимском. Соленость в центральной части Японского моря в сартанское время была на 5–7‰ меньше современной и составляла 27–29‰.
Общее похолодание и сокращение притока теплой воды привело к значительному распространению субарктической микрофлоры и микрофауны. Ее южная граница по сравнению с современной продвинулась на юг на 800–1000 км. В южной части Японского моря содержание субарктических видов составляло 40–60%. Исключением стал район юго-западной части моря, где обитала бореальная фауна. Холодное Приморское палеотечение совершало циклонический круговорот и охватывало почти всю акваторию Японского моря.
Ослабление общей циркуляции морских вод при одновременном усилении холодного Приморского течения на фоне такого похолодания климата привело к сильной ледовитости Японского моря. Влиянием ледового режима определило хорошо выраженную меридиональную асимметрию гольцовой зоны, развитие на побережье Японского моря ландшафта лесотундр и многолетней мерзлоты.
Столь далекое проникновение к югу (до 40° с. ш.) многолетней мерзлоты вдоль побережья Японского моря было связано, помимо общерегионального похолодания, еще и с развитием ледового покрова на поверхности моря. Этим объясняется широкое распространение в Японском море неритическо-сублиторальной группы диатомей, появление бентосных форм ( Achnanthes taeniata, Bacterosira fragilis, Parosira glacialis) и ослабление роли океанического планктона. Ледовитость Японского моря в эпоху наибольшего позднеплейстоценового похолодания была сравнима с современной в Беринговом море.

В северной части Японского моря, особенно в Татарском проливе, сложились благоприятные условия для формирования аккумулятивных берегов. Этому способствовало увеличение общей массы рыхлого материала, выносимого реками и образующегося в результате абразии слабо литифицированных более древних морских осадков, а также общая выровненность побережья и развитие вдоль береговых потоков наносов. Абразионные берега были распространены преимущественно на участках узкого шельфа — у берегов Кореи, в отдельных местах побережья Приморья и Японских островов.
В осадконакоплении ведущая роль принадлежала терригенному материалу, размерность которого обычно уменьшается по мере увеличения глубин водоема. Но в результате ледового разноса крупнообломочный материал (гравий, галька) и пресноводные формы диатомей попадали и в центральную часть Японского моря. Поверхностные течения также перераспределяли осадки. В восточной части Корейского пролива они обусловили распределение тонкозернистого песка, а в западной части — грубозернистого.